WheatBP NEW
CerealsDB home WheatBP home FAQs Contact us
Wheat evolution Wheat development Wheat farming Milling Baking and brewing Wheat research SNPs SNP discovery
Our publications Related Web-sites
Evolution of Wheat
Wheat and other cereals (e.g., rice, barley and maize) belong to the grass family Poaceae (formerly known as the Gramineae) which, with about 10,000 species, represents one of the largest families of flowering plants. The earliest appearence of grasses in evolutionary history has been inferred from fossil records. Pollen possesses two characteristics that make it particularly useful for studying plant evolution: i) it is very resistant to decomposition, and so will often be found in ancient soils; 2) pollen is very distinctive between plant families and species - grass pollen can be easily recognised in mixed pollen samples becuase it is roughly spherical in form and has a single pore on its surface (see main image and Figure 1a). The earliest finds of fossilised grass-like pollen date to between 60 and 70 million years ago (E.A. Kellogg, 2001, Evolutionary History of the Grasses) and so palynologists (scientists who study pollen) place the appearence of the grasses to this period. Other studies, however, indicate that the grasses might be more ancient still. Examining the contents of fossilised dung of plant-eating dinosaurs, scientist have found it to contain types of cells that are only found in the epidermis of grass leaves (figure 1b). Thus, grasses might have originated as much as 80 million years ago and, presumably, dinosaurs dined on grass as reported in the article Dinosaur coprolites and the early evolution of grasses and grazers by Vandana Prasad and co-workers.
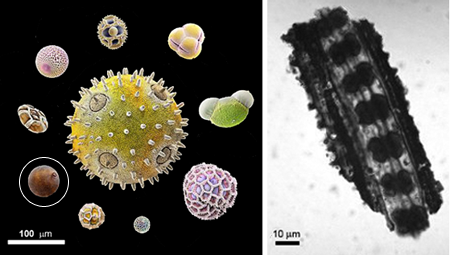
Figure 1. a) Artificially coloured, electron microscope images of various pollen grains to show their distinctiveness and variety of size and form - grass pollen is circled. b) Fossilised grass-epidermal cells found in the dung of plant-eating dinosaurs.
The study of the evolutionary relationship between organisms is called phylogenetics. Traditionally, these relationships would have been inferred from morphological similarities between organisms. However, with the advent of molecular biology, the analysis of DNA and protein sequences have become increasingly important in determining evolutionary relationships. The important characteristic of these molecules in this regard is that, over the course of many generations, their sequence slowly changes due to the accumulation of mutations (see SNP section). Thus, the more distantly related that two species are, the greater the number of differences one sees when comparing any given homologous sequence. If one assumes that mutations occur at a relatively constant rate, these differences can be used as a molecular clock with which to measure evolutionary time. Using such phylogenetic approaches it has been deduced that wheat shared a common ancestor with rice approximately 40 to 54 million years BC. More distantly in time, between 45 and 60 million years BC, there would have been a common ancestor of wheat/rice and sorghum (see phylogenetic tree in figure 2).
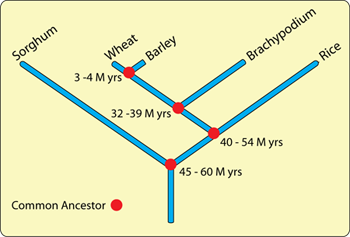
Figure 2. Phylogenetic tree showing the evolutionary relationship between some of the major cereal grasses. Brachypodium is a small grass species that is often used in genetic studies because of its small and relatively simple genome.
The evolution of bread wheat, however, has not been dependent solely on the accumulation of mutations; hybrididation has also been a major factor An understanding of the evolution of wheat has been slowly arrived at through the piecing together of evidence from many different sudies: botany, genetics, archeology and a knowledge of the present distribution of the various grass species thought to have contributed to its evolution. However, cytogenetics (the study of the chromosome complement) has made a major contribution to our present understanding. Breadwheat, Triticum aestivum, is an allopolyploid produced from two separate hybridisation events (see figure 3). Each hybridisation was followed by chromosome doubling in the new hybrid; this enables normal bivalent formation at meiosis and thus the production of fertile plants. The initial hybridisation, that occurred approximately 10,000 years ago, is believed to have been between the two grass species T. urartu (the A genome donor), and T. speltoides (the B genome donor). This new species would have been tetraploid (four complete genome complements) Hexaploid wheat arose as a result of a second hybridisation between the new tetraploid and a third diploid species, T. tauschii (the D genome donor). Again, chromosome doubling must have occurred in order to produce a fertile individual. This new species would then have have 42 chromosomes; that is, six complete genomes each of 7 chromosomes.
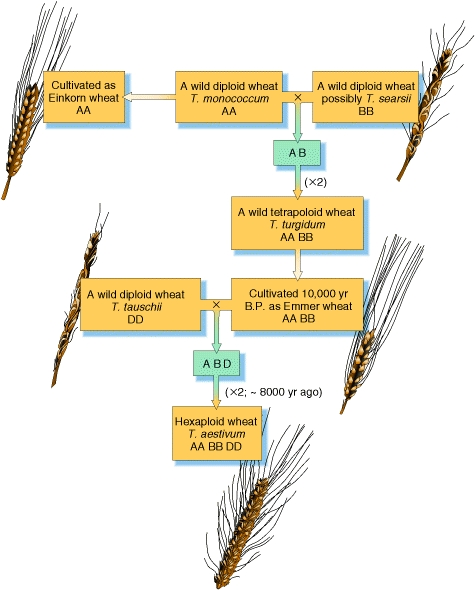
Figure 3. Hybridisations events involved in the evolution of bread wheat, Triticum aestivum. The 'X2' refers to the doubling of the chromosome complement which gives rise in fertile hybrids.
Human intervention must also have played an important role in the evolution of the cultivated wheats and would have been associated with the neolithic shift from an economy based on hunting and gathering to a system based on food production through the domestication of plants and animals. For example, forms of wheat would have been hulled; that is their grains would have been tightly invested by tough paleae and glumes. Early agriculturalists would have selected for varieties with loose paleae and glumes (free threshing varieties) since these would have made the flour making process easier - it goes without saying that these neolithic farmers would not have known that they were selecting for genes that confir this characteristic of the mature plants.
Cultivated Wheat species
Cultivated wheats naturally fall into three main groups on the basis of their genome complement: diploids (possess two copies of the genome and so, in this sense, are like humans), tetraploids (possess four copies of the genome) and hexaploid (six copies). Their assumed centre of origin is the Fertile Crescent, a region extending from south-western Iran, through the Tigris and Euphrates basins in northern Iraq and south-eastern Turkey extending to central Israel and Jordan. Einkorn wheat (one-grained wheat), Triticum monococcum, is believed to be the most ancient cultivated species of wheat, and may be the species from which all cultivated wheat is descended. Studies suggest that einkorn wheat was cultivated in Southeastern Turkey from 10 and 40 thousand years ago. Einkorn wheat is of particular interest because the gluten proteins it produces appear to be less allogenic compared with those produced by bread and durum wheat, thus making it a possible alternative for sufferers of coeliac disease.
Emmer wheat, T. dicoccum, is also very ancient and evidence for its domestication has been found in the Jordan valley and the Damascus basin from about 10 thousand years ago. Later evidence for cultivation has been found in Turkey and Iran. Durum wheat, T. turgidum, is valued for its high gluten content and widely used for pasta, bulgar, couscous and some bread flours. Semolina is coarsely ground durum wheat. Durum wheat is a distinct species, expressing a phenomenon known as tetraploidy, a condition that renders it with 4 copies of each chromosome in its genome. The oldest tetraploid wheat yet discovered comes from the Neolithic of Syria, 8.9 to 8.6 thousand years ago. Durum is an example of a free-threshing or naked wheat, having little or no husk around each grain. By contrast ancient wheat varieties such as einkorn and emmer wheat have very thick husks that make it difficult to extract edible parts of the grain, such as the endosperm and wheatgerm.
Triticum parvicoccum is known to have been cultivated in ancient times and is an early example of a free-threshing wheat. The delicate glumes made it easy to separate the desirable parts of the grain from those that were inedible. Unfortunately T. parvicoccum is now extinct, but is believed to have been tetraploid and may have been the ancestor of hard wheats such as durum wheat.
Spelt flour is derived from a distinct species of wheat that is hexaploid, having 6 copies of each chromosome, T. spelta. Unlike breadwheat, the grain of spelt wheat is not free threshing. It has been suggested that spelt wheat is a hybrid of the extinct tetraploid wheat T. parvicoccum diploid wheat T. tauschii. Spelt wheat has moderately high gluten content, making it suitable for baking bread, and is valued for its distinctive nutty flavour. Spelt wheat flour is high in vitamins and fibre and is now marketed as a health food.